Etapas del ciclo celular donde se encuentran las moléculas reguladoras
Realizado por Mayrin Amarillas
El ciclo celular es un conjunto ordenado de sucesos que conducen al crecimiento de la célula y la división en dos células hijas. Las etapas, son G1-S-G2 y M. El estado G1 quiere decir «GAP 1» (Intervalo 1). El estado S representa la «síntesis», en el que ocurre la replicación del ADN. El estado G2 representa «GAP 2» (Intervalo 2). El estado M representa «la fase M», y agrupa a la mitosis o meiosis (reparto de material genético nuclear) y la citocinesis (división del citoplasma). Las células que se encuentran en el ciclo celular se denominan «proliferantes» y las que se encuentran en fase G0 se llaman células «quiescentes».[1] Todas las células se originan únicamente de otra existente con anterioridad.[2] El ciclo celular se inicia en el instante en que aparece una nueva célula, descendiente de otra que se divide, y termina en el momento en que dicha célula, por división subsiguiente, origina dos nuevas células hijas.En el artículo de puntos de control del ciclo celular, analizamos el porqué de las transiciones del ciclo celular: los factores que considera una célula cuando decide si avanza o no en el ciclo celular. Estas incluyen señales externas (como las señales moleculares) e internas (como el daño del ADN).
Las señales como estas actúan al cambiar la actividad de los reguladores centrales del ciclo celular dentro de la célula. Estos reguladores centrales del ciclo celular pueden causar eventos clave, tales como la replicación del ADN o la separación de los cromosomas. También se cercioran de que los eventos del ciclo celular ocurran en el orden correcto.
En este artículo, veremos algunos de los más importantes reguladores del ciclo celular: proteínas llamadas ciclinas, enzimas llamadas Cdks y un complejo enzimático llamado APC/C.
Ciclinas
Las ciclinas están entre los reguladores centrales más importantes del ciclo celular. Las ciclinas son un grupo de proteínas relacionadas, y en seres humanos y la mayoría de los demás eucariontes existen cuatro tipos básicos: ciclinas de G
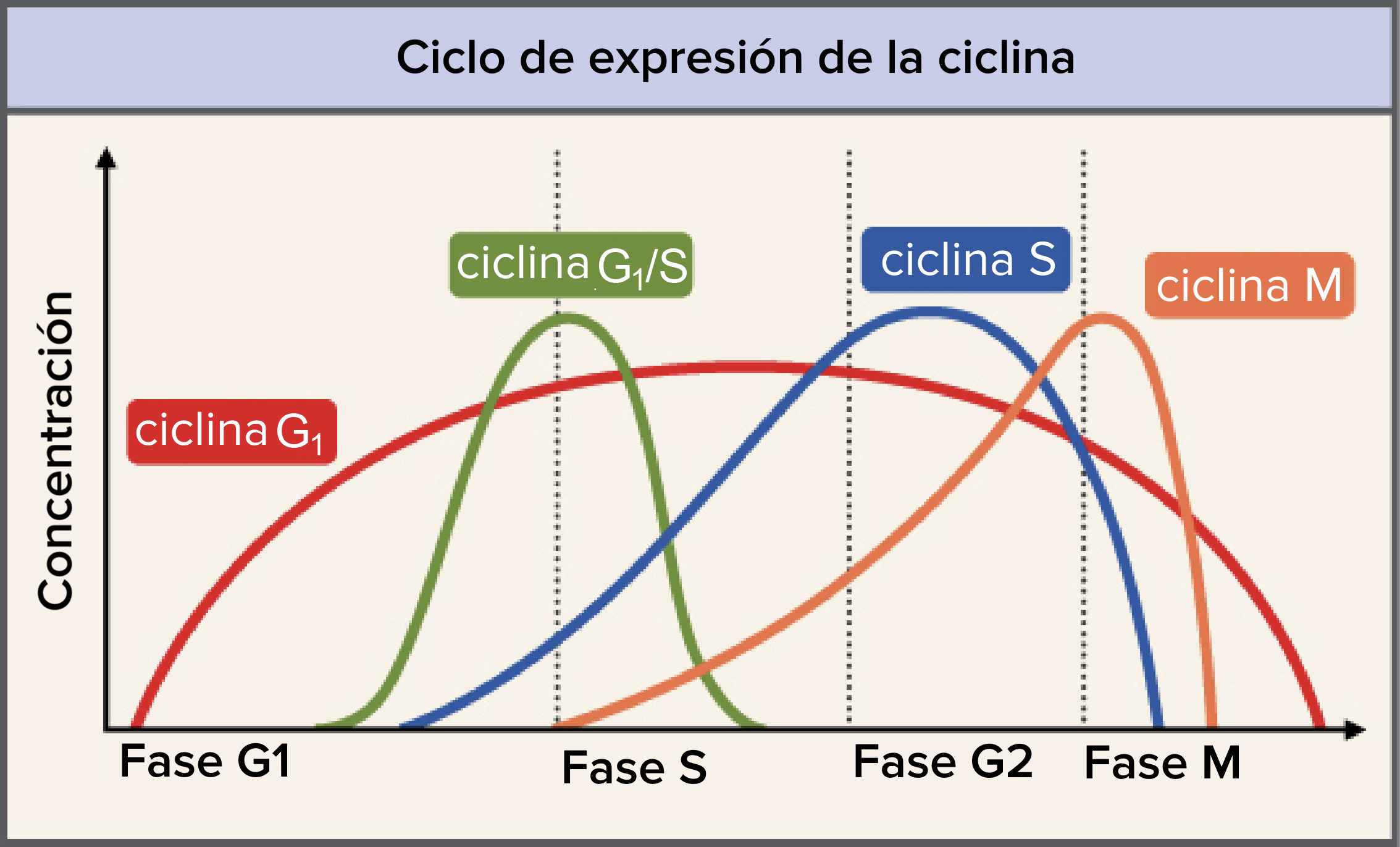
Los niveles de las diferentes ciclinas varían considerablemente a lo largo del ciclo celular, como se muestra en el diagrama de la derecha. Una ciclina típica se presenta en niveles bajos en la mayor parte del ciclo, pero aumenta intensamente en la fase donde es necesaria. La ciclina M, por ejemplo, alcanza dramáticamente su pico en la transición de la fase G
son diferentes en que son necesarias durante la mayor parte del ciclo celular.
Cinasas dependientes de ciclinas
Para promover el ciclo celular, una ciclina debe activar o inactivar muchas proteínas blanco en el interior de la célula. Las ciclinas dirigen los acontecimientos del ciclo celular mediante la asociación con una familia de enzimas llamadas cinasas dependientes de ciclina (Cdks). Una Cdk solitaria es inactiva, pero la unión a una ciclina la activa, la vuelve una enzima funcional y le permite que modifique proteínas blanco.
¿Cómo funciona esto? Las Cdk son cinasas, enzimas que fosforilan (unen a grupos fosfatos) proteínas blanco específicas. La unión del grupo fosfato actúa como un interruptor y hace a la proteína más o menos activa. Cuando una ciclina se une a Cdk, tiene dos efectos importantes: activa la Cdk como una cinasa, pero también dirige a la Cdk a un conjunto específico de proteínas blanco, las apropiadas para el periodo del ciclo celular controlado por la ciclina. Tal sería el caso de las ciclinas G

En general, los niveles de Cdk se mantienen relativamente constantes a lo largo del ciclo celular, pero la actividad de Cdk y las proteínas blanco cambian a medida que los niveles de las diversas ciclinas suben y bajan. Además de necesitar una compañera ciclina, las Cdk también deben fosforilarse en un sitio en particular para estar activas (no se muestra en los diagramas de este artículo) y también pueden regularse negativamente mediante la fosforilación de otros sitios
.Las ciclinas y las Cdk están muy conservadas evolutivamente, lo que significa que se encuentran en muchos tipos diferentes de especies, desde las levaduras hasta las ranas y los humanos. Los detalles del sistema varían un poco: por ejemplo, las levaduras tienen solo una Cdk, mientras que los humanos y otros mamíferos tienen varias Cdks que se utilizan en diferentes fases del ciclo celular. (Sí, esta es una pequeña excepción a la regla "los niveles de las Cdks no cambian"). Pero los principios básicos son muy similares, así que las Cdks y los diferentes tipos de ciclinas pueden encontrarse en cada especie.
Factor promotor de la maduración (MPF)
Un ejemplo famoso de cómo las ciclinas y las Cdks trabajan juntas para controlar la transición del ciclo celular es el del factor promotor de la maduración (MPF). El nombre se remonta a la década de 1970, cuando los investigadores encontraron que las células en la fase M contenían un factor desconocido que podría forzar a los óvulos de la rana (atoradas en la fase G)
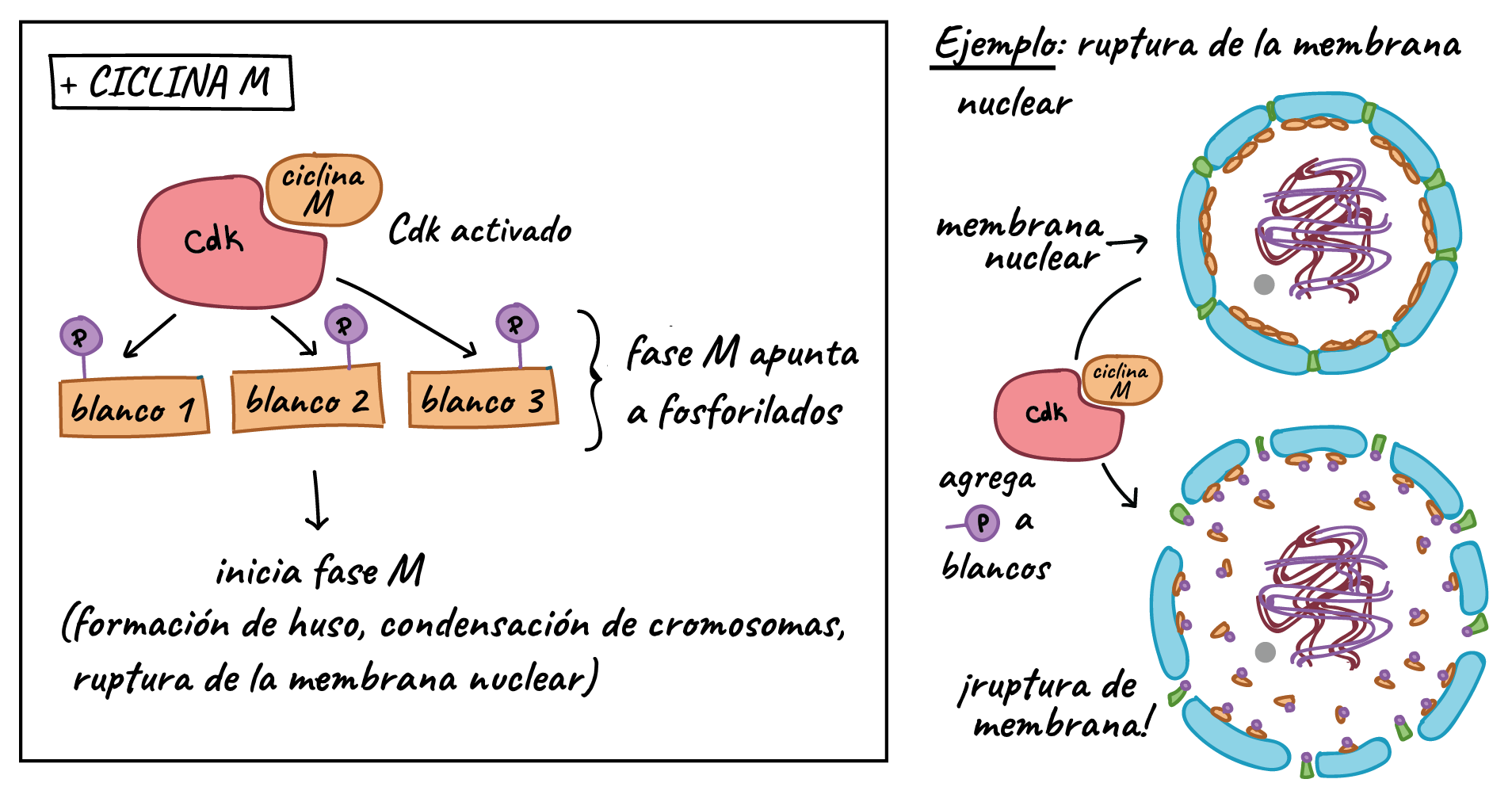
MPF es un buen ejemplo de cómo las ciclinas y Cdks pueden trabajar juntas para promover una transición del ciclo celular. Igual que una ciclina típica, la ciclina M se mantiene en niveles bajos durante la mayoría del ciclo celular, pero se acumula a medida que la célula se aproxima a la transición G
El complejo promotor de la anafase/ciclosoma (APC/C)
Además de inducir los eventos de la fase M, el MPF también provoca su propia destrucción mediante la activación del complejo promotor de la anafase/ciclosoma (APC/C), un complejo proteico que causa que las ciclinas M se destruyan a partir de la anafase. La descomposición de las ciclinas M expulsa a la célula de la mitosis y permite que las nuevas células hijas entren a G
¿Cómo hace su trabajo el APC/C? Como una Cdk, el APC/C es una enzima, pero tiene un tipo de función diferente al de una Cdk. En lugar de unir un grupo fosfato a su objetivo, se añade un pequeña etiqueta proteica llamada ubiquitina (Ub). Cuando un blanco es marcado con ubiquitina, se envía al proteasoma, que puede considerarse como la papelera de reciclaje de la célula, y se destruye. Por ejemplo, el APC/C adhiere una etiqueta de ubiquitina a las ciclinas M, lo que causa que estas sean degradadas por el proteasoma y permite que las células hijas recién formadas entren en la fase G
El APC/C también utiliza etiquetas de ubiquitina para provocar la separación de las cromátidas hermanas durante la mitosis. Si el APC/C recibe las señales adecuadas en la metafase, pone en marcha una cadena de eventos que destruye la cohesina, el adhesivo proteico que mantiene juntas a las cromátidas hermanas
- El APC/C primero adiciona una etiqueta de ubiquitina a una proteína llamada segurina, que la envía a reciclaje. La segurina normalmente se une e inactiva una proteína llamada separasa.
- Cuando la segurina es enviada a reciclaje, la separasa se activa y puede hacer su trabajo. La separasa disgrega la cohesina que mantiene unidas a las cromátidas hermanas y les permite separarse.

Puntos de control y reguladores
Las Cdk, ciclinas y APC/C son reguladores directos de las transiciones del ciclo celular, pero no siempre están al mando. Por el contrario, responden a las señales que provienen de dentro y fuera de la célula. Estas señales influyen sobre la actividad de los reguladores centrales para determinar si la célula avanza en el ciclo celular. Las señales positivas, como los factores de crecimiento, típicamente aumentan la actividad de las Cdk y ciclinas, mientras que las negativas, como el daño al ADN, generalmente disminuyen o bloquean la actividad.
A modo de ejemplo, veamos cómo el daño del ADN detiene el ciclo celular en G
El daño del ADN puede suceder, y sucederá, en muchas células del cuerpo humano durante
la vida de una persona (por ejemplo, debido a los rayos UV del sol).
Las células deben ser capaces de hacer frente a este daño, reparándolo si es posible y previniendo
la división celular si no es posible.
Una proteína llamada p53 es clave en la respuesta al daño del ADN, es un famoso supresor
tumoral a menudo descrito como "el guardián del genoma".
la vida de una persona (por ejemplo, debido a los rayos UV del sol).
Las células deben ser capaces de hacer frente a este daño, reparándolo si es posible y previniendo
la división celular si no es posible.
Una proteína llamada p53 es clave en la respuesta al daño del ADN, es un famoso supresor
tumoral a menudo descrito como "el guardián del genoma".
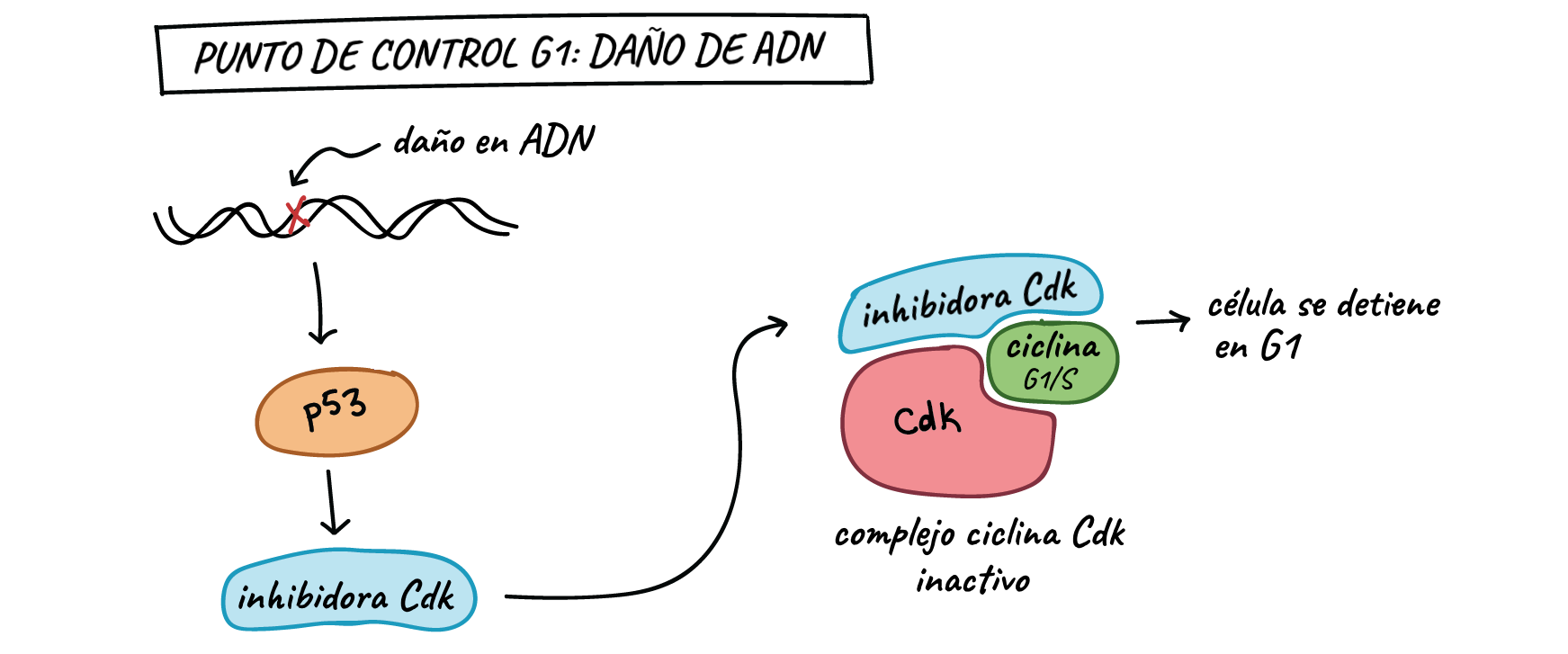
Primero, detiene el ciclo celular en el punto de control G
Al activar la producción de las proteínas inhibidoras de Cdk (CKI). Las proteínas CKI se
fijan a los complejos Cdk-ciclina y bloquean su actividad (ve el diagrama siguiente), ganando
tiempo para la reparación del ADN. El segundo trabajo de p53 es activar las enzimas de
reparación del ADN. Si el daño al ADN no es reparable, p53 desempeñará su tercer y último
papel: activar la muerte celular programada para que el ADN dañado no sea transmitido.
fijan a los complejos Cdk-ciclina y bloquean su actividad (ve el diagrama siguiente), ganando
tiempo para la reparación del ADN. El segundo trabajo de p53 es activar las enzimas de
reparación del ADN. Si el daño al ADN no es reparable, p53 desempeñará su tercer y último
papel: activar la muerte celular programada para que el ADN dañado no sea transmitido.
Al asegurar que las células no se dividen cuando su ADN está dañado, p53 previene que las mutaciones (cambios en el ADN) se transmitan a las células hijas. Cuando p53 está defectuoso o ausente, las mutaciones se acumulan rápidamente, lo que puede conducir al cáncer. De hecho, de todo el genoma humano completo, p53 es el gen que está mutado con mayor frecuencia en los cánceres.

Comentarios
Publicar un comentario